

 Blog SVT - Lycée Vauban 15 rue
Bousy 08600 Givet
Blog SVT - Lycée Vauban 15 rue
Bousy 08600 Givet
 Par Lydie.
Par Lydie.
La trisomie 21
Le syndrome de Down, aussi appelé trisomie 21, est une maladie chromosomique congénitale provoquée par la présence d'un chromosome surnuméraire pour la 21e paire. Ses signes cliniques sont très nets, on observe un retard cognitif, associé à des modifications morphologiques particulières.
La maladie a été décrite par le médecin britannique John Langdon Down dans un article de 1866. Il est l'auteur des mots mongolien et mongolisme, proposés en référence aux yeux bridés des sujets atteints par le syndrome (l'emploi de ces termes est aujourd'hui officiellement déconseillé).
L'origine chromosomique de cette maladie a été découverte par Jérôme Lejeune, médecin français, en juillet 1958. Ce fut la première maladie pour laquelle fut mise en évidence la relation entre le génotype et le phénotype. Il lui donna le nom de « trisomie 21 » ce qui se décompose en « tri » voulant dire « trois », « some » voulant dire « chromosome », c’est-à-dire « trois chromosomes 21 ».
En savoir plus...
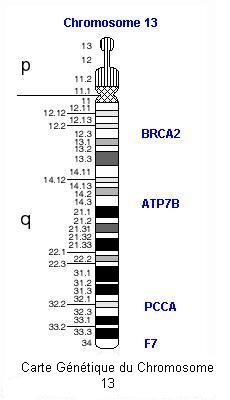
Voir un caryotype standard : homme - femme.
Voir un caryotype de trisomie 21.
Propositions d'activités relatives au diagnostic et au dépistage de la trisomie 21 (Académie de Versailles). Page très intéressante pour les élèves de TS, je vous conseille de visiter les liens pour accéder à une documentation riche.
Dépistage de la trisomie 21, un cours niveau TS
Dépistage de la trisomie 21 : La clarté nucale.
Dépistage par les marqueurs sériques (hcg et afp).
Les trisomies autosomiques
En savoir plus sur les trisomies autosomiques (quelques exemples) :
La trisomie 6.
La trisomie 7.
La trisomie 8.
la trisomie 9 = Syndrome de Warkany
La trisdomie 13 = syndrome de Patau.
La trisomie 18 = syndrome d'Edward.
Les trisomies hétérosomiques
Certaines trisomies atteignent les hétérosomes (chromosomes sexuels), X, ou Y :
Trisomie X (Syndrome Triple X) : l'individu possède trois chromosomes X, le chromosome supplémentaire est toujours donné par la mère, le risque augmente avec l'âge. L'enfant est de sexe féminin. Une fille sur mille est atteinte de cette maladie génétique.
Le Syndrome de Klinefelter : L'individu possède deux chromosomes X et un chromosome Y (XXY). L'enfant est de sexe masculin — dans la plupart des cas, mais peut aussi être de sexe féminin ou indéfini (malformations probables).
Le Syndrome de Jacob : l'individu possède un chromosome Y en double exemplaire, et un chromosome X (XYY). Le chromosome Y supplémentaire est dû à un problème au cours de la spermatogénèse chez le père. Environ un homme sur 850 développe au moins un spermatozoïde double Y. L'enfant est de sexe masculin.
Le dépistage des anomalies chromosomiques
Une technique de dépistage des anomalies chromosomiques : la FISH.



















{kind=link}